Dr. Adrián Ponce-Alvarez
Theoretical neuroscientist
Ramón y Cajal Research Fellow at
Department of Mathematics
Dynamical Systems Group
Polytechnic University of Catalonia, Spain.


About me
Selected publications
Grabenhorst F, Ponce-Alvarez A, Battaglia-Mayer A, Deco G, Schultz W (2023) A view-based decision mechanism for rewards in the primate amygdala. Neuron 111, 1-14.
Ponce-Alvarez A, Kringelbach ML, Deco G (2023) Critical scaling of whole-brain resting-state dynamics. Nature Communications Biology 6, 627.
Ponce-Alvarez A, Uhrig L, Deco N, Signorelli CM, Kringelbach ML, Jarraya B, Deco G (2022) Macroscopic quantities of collective brain activity during wakefulness and anesthesia. Cerebral Cortex 32(2), 298–311
Pfeffer T, Ponce-Alvarez A, Meindertsma T, Gahnström C, van den Brink RL, Nolte G, Tsetsos K, Engel AK, Deco G, Donner TH (2021) Circuit mechanisms for the chemical modulation of cortex-wide network interactions and exploration behavior. Science Advances, 7(29), eabf5620.
Ponce-Alvarez A, Mochol G, Hermoso-Mendizabal A, de la Rocha J, Deco G (2020) Cortical state transitions and stimulus response evolve along stiff and sloppy parameter dimensions, respectively. eLife 9, e53268.
Hahn G, Ponce-Alvarez A, Deco G, Aertsen A, Kumar A (2019) Portraits of communication in neuronal networks. Nature Reviews Neuroscience 20, 117-127.
Ponce-Alvarez A*, Jouary A*, Privat M, Deco G, Sumbre G (2018) Whole-brain neuronal activity displays crackling noise dynamics. Neuron 100, 1446-1459.
Ponce-Alvarez A, Deco G, Hagmann P, Romani GL, Mantini D, Corbetta M (2015a) Resting-state temporal synchronization networks emerge from connectivity topology and heterogeneity. PLoS Computational Biology e1004100.
Ponce-Alvarez A*, Thiele A*, Albright TD, Stoner GR, Deco G (2013) Stimulus-dependent variability and noise correlations in cortical MT neurons. PNAS 110(32):13162-7.
Deco G*, Ponce-Alvarez A*, Mantini D, Romani GL, Hagmann P, Corbetta M (2013) Resting-state functional connectivity emerges from structurally and dynamically shaped slow linear fluctuations. Journal of Neuroscience 33(27):11239-52.
I am a theoretical neuroscientist employed as a Ramón y Cajal Research fellow at the Polytechnic University of Catalonia, Barcelona, Spain.
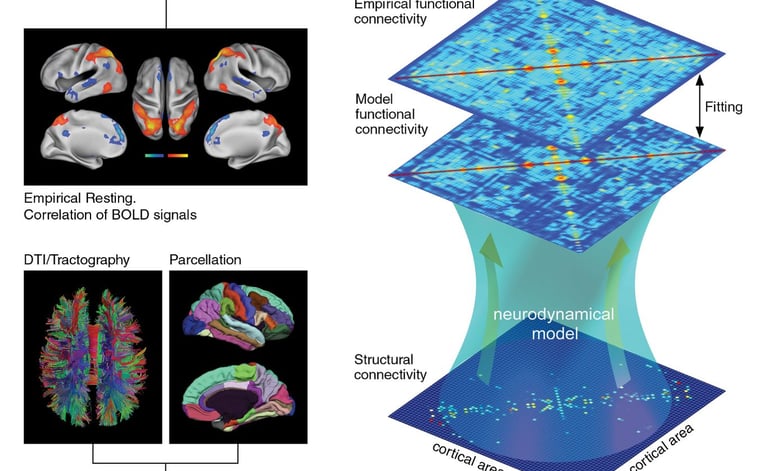
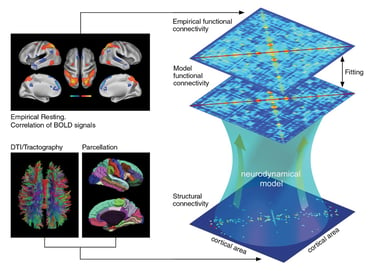
My research concerns the study of neural networks, at different scales and in different states. I focus primarily on how cognitive processes are represented in the spatiotemporal patterns of neuronal activity; how brain regions interact; how collective behaviors emerge in neural networks; and how these collective behaviors underlie brain function/dysfunction and different brain states (e.g., conscious wakefulness, sleep, anesthesia).
My approach uses a combination of data analysis and theory, by means of stochastic dynamical systems, information theory, statistical mechanics, machine learning, and network theory.
I am committed in favour of gender, race, and class equity in Academia. I was member of the Center for Gender Studies (CEdGE) at the Pompeu Fabra University from 2015-2022.





CONTACT
Email: adrian.ponce@upc.edu
Polytechnic University of catalonia (UPC)
Department of Mathematics
Dynamical Systems Group